¡Descarga Inmunidad Innata: Mecanismos y Células y más Resúmenes en PDF de Inmunología solo en Docsity!
FUNCIONES:
- Limita la proliferación de los microorganismos con los que nos enfrentamos.
- Elimina infecciones ocasionadas por microorganismos patógenos o las contiene hasta que se desarrolla la respuesta inmune adaptativa. Consta de las defensas contra la infección que aún antes del ataque de un patógeno están listas para activarse de inmediato. Incluye barreras:
- Físicas:
- PIEL
- MEMBRANAS MUCOSAS
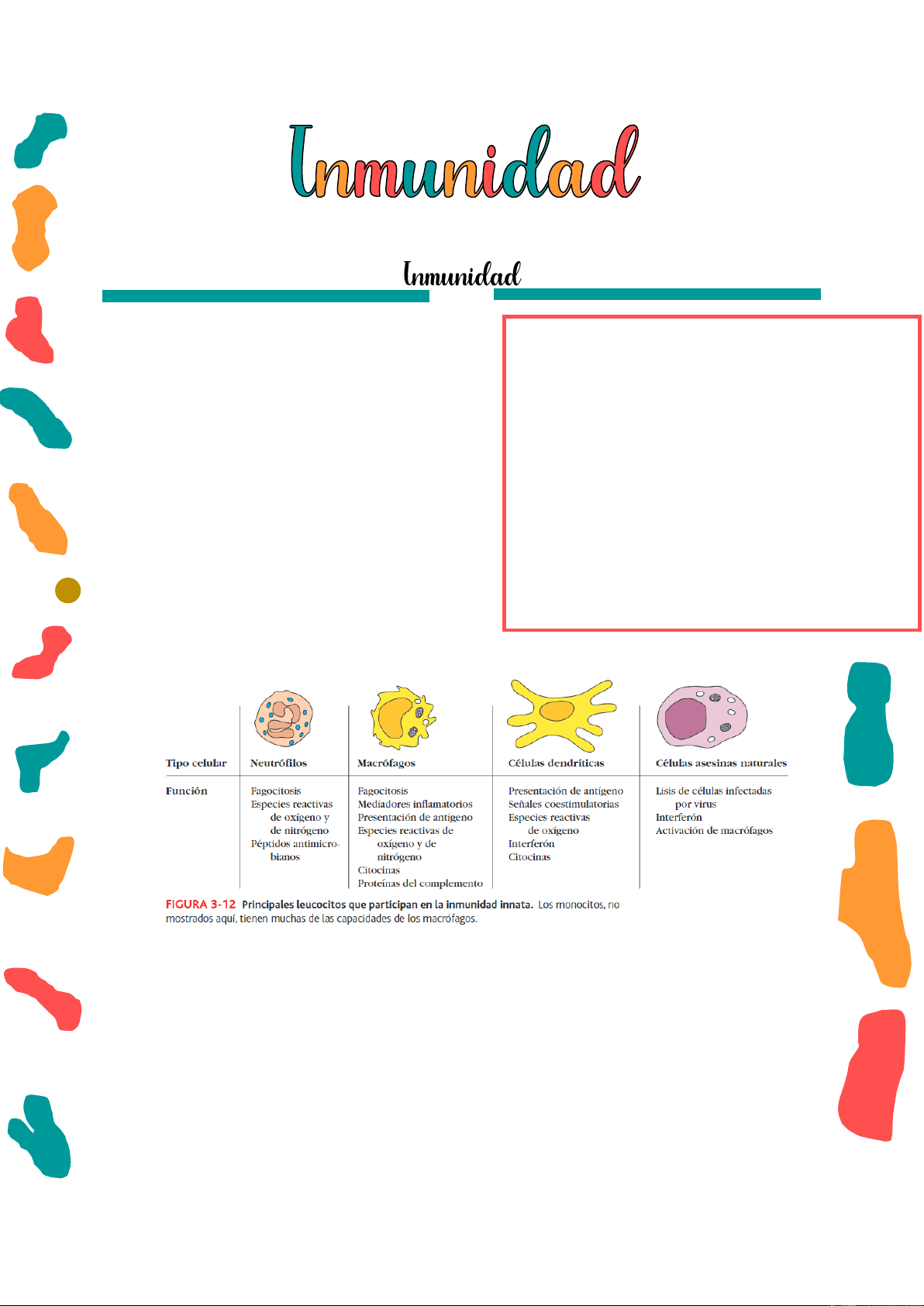
- Químicas: Innata Células que median la inmunidad innata:
- Granulocitos: neutrófilos, eosinófilos y basófilos
- Mastocitos
- Células NK
- Células NKT
- Células dendríticas convencionales
- Células dendríticas plasmocitoides
- Células epiteliales
- Células endoteliales
- Células parenquimatosas
- ACIDEZ DEL CONTENIDO ESTOMACAL
- MOLECULAS SOLUBLES CON ACTIVIDAD
ANTIMICROBIANA
- Celulares:
- CÉLULAS CON RECEPTORES SENSIBLES QUE DETECTAN PRODUCTOS MICROBIANOS E INSTIGAN UN CONTRAATAQUE. La respuesta a la invasión por un microorganismo infeccioso que supera las barreras iniciales de la piel y membranas mucosas es rápida; típicamente se inicia a los pocos minutos de la invasión. Es posible que algunos patógenos logren evadir estas defensas. Por ello existe la INMUNIDAD ADAPTATIVA o ADQUIRIDA. BARRERAS ANATÓMICAS
- PIEL
- MEMBRANAS MUCOSOS: incluye epitelios mucosos que recubren las vías respiratorias, digestivas y urogenitales y aíslan el interior del cuerpo contra los patógenos del mundo exterior. MECANISMOS INMUNEN QUE PREVIENEN EL ESTABLECIMIENTO DE UN FOCO INFECCIOSO EN MUCOSAS:
- Secreciones mucosas (moco): dificultan el acceso de patógenos al epitelio, bloquean moléculas empleadas por los patógenos para adherirse al epitelio.
- Factores químicos presentes en secreciones mucosas: péptidos antimicrobianos, lisozima, lactoferrina, etc.
- Factores físicos: descamación (evita adherencia), oscilaciones ciliares, movimientos peristálticos
Estos sensores moleculares reconocen motivos estructurales generales con alto grado de conservación dentro de una especie microbiana, pero comúnmente están ausentes en el hospedados. Dado que reconocen patrones moleculares generales específicos, se denominan RECEPTORES DE RECONOCIMIENTO DE PATRÓN (PRR), y cuando los patrones se detectan en los patógenos, se les denomina PATRONES MOELCULARES RELACIONADOS CON EL PATÓGENO (PAMP). Entre los PAMP reconocidos por PRR se incluyen combinaciones de azúcares, determinadas proteínas, moléculas portadoras de lípidos específicos y algunos motivos (estructuras repetitivas) de ácidos nucleicos. La restricción del reconocimiento innato a patrones moleculares presentes en los microorganismos hace que el sistema innato se concentre en entidades que pueden causar infección más que en sustancias que son ajenas. Los anticuerpos y receptores de célula T, los sensores de la inmunidad adaptativa, reconocen detalles más finos de la estructura molecular y son capaces de discriminar con especificidad entre antígenos que presentan sólo diferencias estructurales ligeras. La capacidad de los PRR de distinguir entre lo propio y lo extraño es a pruebas de errores, Los PAMP son moléculas, un conjunto diferenciado de componentes microbianos que les permiten a las células de la inmunidad innata el rápido reconocimiento de un proceso infeccioso y la inmediata puesta en marcha de una respuesta que integra la producción de:
- Sustancias con actividad microbiostática o microbicida
- Citocinas
- Quimiocinas Están presentes en los microorganismos, comparten 3 propiedades:
- Se expresan en microorganismos, pero no en el hospedador
- Son compartidos por diferentes microorganismos
- Son esenciales para la supervivencia o la patogenicidad de los microorganismos. Entre los mejor caracterizados:
- Lipopolisacárido (LPS)
- Flagelina
- Peptidoglucano
- Ácidos nucleicos microbianos
porque el patrón molecular al que se dirige el receptor solo es producido por el patógeno y nunca por el hospedador. En la inmunidad innata, la detección de patrones moleculares relacionados con patógeno realizada por mediadores solubles y unidos a membrana pone en juego múltiples componentes inmunitarios. Entre los mediadores solubles se incluyen iniciadores del SISTEMA DEL COMPLEMENTO, como LECTINA DE UNIÓN A MANOSA (MBL), y PROTEÍNA C REACTIVA (CRP). Si el patógeno porta PAMP que son reconocidos por estos mediadores, se activará el sistema del complemento. Una parte de dicho sistema es un grupo de proteínas, que cuando se activan, forman agregados los cuales hacen agujeros en las membranas celulares de los microorganismos contra los que se dirigen, a los que dan muerte por lisis. El sistema de complemento también incluye glucoproteínas séricas, que cuando son activadas, promueven la captación de microorganismos por fagocitos (OPSONIZACIÓN). El sistema del complemento ocupa una posición intermedia entre los sistemas inmunitarios innato y adaptativo: la cascada del complemento, que desemboca en la opsonización o lisis de los invasores, puede ser activada por moléculas que reconocer PAMP (inmunidad innata) o por anticuerpos (inmunidad adaptativa) que se unen a antígenos extraños específicos. Además, algunos de los subproductos promueven la inflamación y por tanto llevan leucocitos al sitio de infección, con lo que lanzan otro nivel de respuesta. Células dendríticas inmaduras y macrófagos del tejido invadido tienen diversos receptores, entre ellos el grupo más importante de receptores innatos: receptores Toll (TLR), que detectan productos microbianos. Cada TLR reacciona con un producto microbiano específicos. Estos hacen posible que las
inflamación y ayudan a dirigir la inmunorreacción adaptativa del hospedador. Todas las funciones inmunitarias innatas ocurren en una fase temprana de la infección. Las citocinas liberadas de células que participan en la respuesta innata modifican la naturaleza de las inmunoreacciones adaptativas ulteriores a la infección. INFLAMACIÓN Cuando los patógenos superan las barreras externas de la inmunidad innata- piel y mucosas-, la infección o lesión tisular resultante puede inducir una compleja cascada de fenómenos conocida como REACCIÓN INFLAMATORIA. La inflamación puede ser aguda, o crónica. La reacción inflamatoria aguda combate las primeras fases de una infección y pone en marcha procesos que llevan a la reparación del tejido dañado. En pocas horas, en la región inflamada se adhieren leucocitos a las células endoteliales y atraviesan las paredes de los capilares para ingresar en los espacios tisulares, un proceso llamada EXTRAVASACIÓN. Estos leucocitos fagocitan patógenos invasores y liberan mediadores moleculares que contribuyen a la reacción
inflamatoria y al reclutamiento y activación de células efectoras. Entre los mediadores se encuentran proteínas reguladoras de bajo peso molecular de la familia de citocinas. Las citocinas son secretadas por glóbulos blancos y otras células del cuerpo en respuesta a estímulos, y tienen papeles importantes en la regulación del desarrollo y el comportamiento de las células efectoras inmunitarias.
- QUIMIOCINAS: subconjunto de citocinas, cuya característica distintiva es la capacidad de actuar como QUIMIOATRACTANTES (agentes que hacen que las células se desplacen hacia los sitios en que esos agentes se encuentran en mayores concentraciones). No todos los quimioatrayentes son quimicinas. Otros quimioatrayentes importantes son los subproductos del complemento (C5a y C3a) y diversos pérptidos N-formilo producidos por la degradación de proteínas bacterianas durante una infección. La unión de quimiocinas u otros quimioatrayentes a receptores en la membrana de las células neutrofílicas desencadena una señal activadora que induce un cambio conformacional en una molécula de la membrana del neutrófilo llamada INTEGRINA, lo que incrementa su afinidad por MOLÉCULAS DE ADHESIÓN INTERCELULAR (ICAM) presentes en el endotelio. Las quimiocinas son los reguladores más importantes y versátiles del tráfico de LEUCOTIOS, ya que controlan de manera selectiva la adhesión, quimiotaxis y activación de diversas subpoblaciones leucocitarias. Las quimiocinas inflamatorias suelen ser inducidas en respuesta a infección o a CITONAS PROINFLAMATORIAS. Las quimiocinas hacen que los leucocitos ingresen en diversos sitios tisulares al
fuerzas cortantes cuando la sangre circulante fluye alrededor de la célula. A medida que las distintas regiones se unen y desprenden, el neutrófila gira dando tumbos sobre la superficie del endotelio. Mientras el neutrófilo rueda sobre el endotelio, es posible que encuentre quimiocinas u otros quimioatrayentes que se han producido en el sitio de proceso inflamatorio. La ulterior interacción entre integrinas y ICAM estabiliza la adhesión del neutrófilo a la celula endotelial, permite abrirse paso entre células del endotelio. b. Activación por estimulo quimioatrayente c. Detención y adhesión d. Migración transendotelial
- LAS PROTEÍNAS DE LA REACCIÓN DE FASE AGUDA CONTRIBUYEN A LA INMUNIDAD INNATA: cambios en la concentración de varias proteínas séricas durante la fase aguda de la enfermedad, la que precede a la recuperación o la muerte. Los cambios séricos en conjunto, REACCIÓN DE FASE AGUDA (APR), y las proteínas cuyas concentraciones aumentan o disminuyen
durante esa fase se conocen como PROTEÍNAS DE REACCIÓN DE FASE AGUA (PROTEINAS APR). La reacción de fase aguda es inducida por señales que viajan por la sangre desde sitios de lesión o infección. El hígado es uno de los principales sitios de síntesis, y las citocinas proinflamatorias TNF-alfa, IL-1 e IL-6 son las principales señales responsable de inducir la reacción. La producción de estas citocinas es una de las respuesta tempranas de los fagocitos, y el aumento en las concentraciones de proteína C reactiva y otras proteínas de fase agua como el complemento contribuye a la defensa de diversas maneras. La proteína C reactiva pertenece a pentraxinas, que fijan ligandos en una reacción dependiente de Ca. Entre los ligandos reconocidos por CRP están un polisacárido y fosforilcolina, que se encuentran en la superficie de muchos microorganismos. La proteína C reactiva unida a estos ligandos en la superficie de un patógeno promueve la captación por fagocitos y activa un ataque mediado por complemento contra el invasos. La lectina de unión a manosa es una proteína de fase aguda que reconoce patrones moleculares que contienen manosa en los microorganismos.
Después de la activación inicial, los diferentes componentes del complemento interactúan en una cascada muy regulada para llevar a cabo varias funciones básicas:
- Lisis de células, bacterias o virus
- Opsonización, que promueve la fagocitosis de antígenos particulados
- Unión a receptores de complemento específicos en células del sistema inmunitario, lo que desencadena funciones características de dichas células, inflamación y secreción de moléculas inmunorreguladoras
- Depuración inmunitaria, elimina complejos inmunitarios de circulación y los deposita en el bazo y el hígado. COMPONENTES: proteínas y glucoproteínas solubles que lo componen se sintetizan en los hepatocitos, también producen cantidades de importancia monocitos sanguíneos, macrófagos tisulares y células epiteliales de los aparatos digestivo y genitourinario. Los componentes se designan con numerales (C1-C9), letras, o nombres comunes. Los fragmentos peptídicos que se forman por activación de un componente se indican con letras pequeñas. En la mayor parte, los fragmentos más pequeños que resultan de la escisión de un componente se designan “a” y el fragmento más grande “b”. Los fragmentos más grandes se
unen al blanco cerca del sitio de activación, y los más pequeños se difunden desde el sitio y pueden precipitar reacciones inflamatorias localizadas por unión a receptores específicos. ACTIVACIÓN DEL COMPLEMENTO Las etapas tempranas, que culminan con la formación de C5b, pueden ocurrir por vías:
- CLÁSICA: inicia con la formación de complejos de antígeno y anticuerpo solubles o con la unión de anticuerpo a antígeno en un blanco conveniente. La igM y subclases de IgG (IgG1, IgG2, IgG3) pueden activarla. Induce cambios de conformación en la porción Fc de la molécula de IgM, los cuales exponen un sitio de unión para el componente C1 del sistema del complemento. C3, se escinde con rapidez en C3a y C3b por la actividad enzimática de la convertasa de C3.
- ALTERNA: genera productos activos similares a los de la vía clásica, pero lo hace sin necesidad de complejos de antígeno y anticuerpo para iniciarse. Es un componente del sistema inmunitario innato porque no requiere anticuerpo. El C3 sérico, sufre hidrólisis espontánea lenta para producir C3a y C3b. El componente C3b pueden unirse a antígenos de superficie extraños o a células del hospedador. La unión de C3b expone un sitio en el factor B que sirve como sustrato para un proteína sérica con actividad enzimática, FACTOR D. Este último escinde el factor B unido a C3b, con lo que se libera un pequeño fragmento y se genera C3bBb. Este complejo posee actividad de
membrana de la célula blanco, permite la libre difusión de iones y moléculas pequeñas a través de la membrana. El resultado final de la activación de las vías es la producción de una convertasa de C5 activa. Las moléculas del sistema inmunitario adaptativo son:
- ANTICUERPO
- RECEPTOR DE LA CÉLULA T Exhiben mayor gardo de especificidad, al reconocer DETERMINANTES ANTIGÉNICOS o EPÍTOPOS ESPECÍFICOS. Los EPÍTOPOS son las regiones con actividad inmunitaria de un inmunógeno que se unen a receptores de membrana específicos de antígeno en los linfocitos o a anticuerpo secretados. Los ANTICUERPOS son proteínas de unión a un epítopo que existen en dos formas:
- Constituyentes unidos a membrana de las células B: confieren especificidad antigénica en las células B
- Moléculas solubles secretadas por células plasmáticas. Todos los anticuerpos comparten características estucturales, se unen a antígeno y participan en funciones efectoras. Los ANTÍGENOS son moléculas que interactúan con el receptor de inmunoglobulina de las células B (o con el receptor de célula T cuando se encuentra en forma de complejo con MHC) ESTRUCTURA BÁSICA Y FUNCIÓN DE LOS ANTICUERPOS El reconocimiento de un inmunógeno por los anticuerpos de adaptativa
superficie de la célula B inicia la proliferación y difereciacion en linfocitos B de memoria y células plasmáticas. Las moléculas de anticuerpo tienen una estructura común de 4 cadenas polipeptídicas. Se integra de:
- 2 cadenas ligeras (L) idénticas
- 2 cadenas pesadas (H) Cada cadena ligera está unida a una pesada por un enlace disulfuro y por interacciones no covalentes. Los dos fragmentos idénticos tenían actividad de unión de antígeno, se denominan FRAGMENTOS Fab. El otro fragmento carecía de actividad de unión del antígeno. Se llamo FRAGMENTO Fc. La mitad amino terminal de la cadena, REGIÓN VARIABLE (V). La mitad carboxilo terminal, REGIÓN CONSTANTE (C) Hay 5 tipos de cadenas principales:
- μ = IgM se produce en respuesta primaria a antígeno, es la primera que sintetiza el recién nacido.
• δ = IgD
- γ = IgG: la más abundante en el suero. 4 subclases (IgG1, IgG2, IgG3, IgG4)
- IgG1, IgG3 e IgG4 cruzan con facilidad la placenta y tienen papel importante en protección del feto en desarrollo.
PRESENTADORAS DE ANTÍGENO (CPA), cuya función es la de presentar péptidos antigénicos a través de moléculas CMH. La porción variable del TCR es específica, no para un péptido, sino para un complejo péptido-molécula del CMH. La diferencia en el reconocimiento antigénico por parte de los linfocitos T y B determina que ciertos epítopos microbianos serán reconocidos solo por los linfocitos B y otros solo por los linfocitos T. La inmunidad adaptativa demora más tiempo porque debe completar:
- activación
- Proliferación
- Diferenciación:
- Células efectoras
- Células memoria. INMUNIDAD MEDIADA POR LT La MADURACIÓN de los genes de TCR de la línea germinal y la expresión de diversos marcos de membrana. En el timo ,las células T en desarrollo, TIMOCITOS, proliferan y se diferencian a lo largo de vías del desarrollo que generan subpoblaciones funcionalmente distintas de células T madera. El timo, el origen principal de las células T, es el sitio en que se diversifican y se configuran en un repertorio eficaz de células T primarias por dos procesos de selección extraordinarios:
- Selección positiva: supervivencia solo de células T cuyos TCR son capaces de reconocer las MHC propias. Creacion de células T restringidas al MHC propia.
- Selección negativa: elimina células T que reaccionan demasiado intensamente con el MHC propio o con MHC propio mas péptidos propios. ACTIVACIÓN DE LA CELULA T es iniciada por la interacción del complejo TCR-CD3 con un péptido antigénico procesado único a una molécula MHC clase I (células CD8+) o clase II (células CD4+) en la superficie de una célula presentadora de antígeno. La interacción de la célula T con antígeno precipita una cascada de fenómenos bioquímicos que inducen a las células T restantes a ingresar al ciclo celular, proliferar y diferenciarse en células de memoria o efectoras. DIFERENCIACIÓN Las células T CD4+ y CD8+ salen del timo y pasan a la circulacion como cleulas en reposo de la etapa G0 del ciclo celular. Hay casi el doble de células T CD4+ en la periferia. Las células T que no encuentran aún antígeno (CÉLULAS T VÍRGENES) se caracterizan por cromatina condensada, poco citoplasma y escasa actividad transcripcional. Las células vírgenes T circulan de modo continuo entre los sistemas sanguíneo y linfático. Durante la recirculación, las células T vírgenes residen en tejidos linfoides secundarios, como los ganglios linfáticos. Si una célula virgen no encuentra antígeno en un ganglio linfático, sale a través de los linfáticos efentes que drenan al final en el conductos torácico y pasa de nueva cuenta a la sangre.